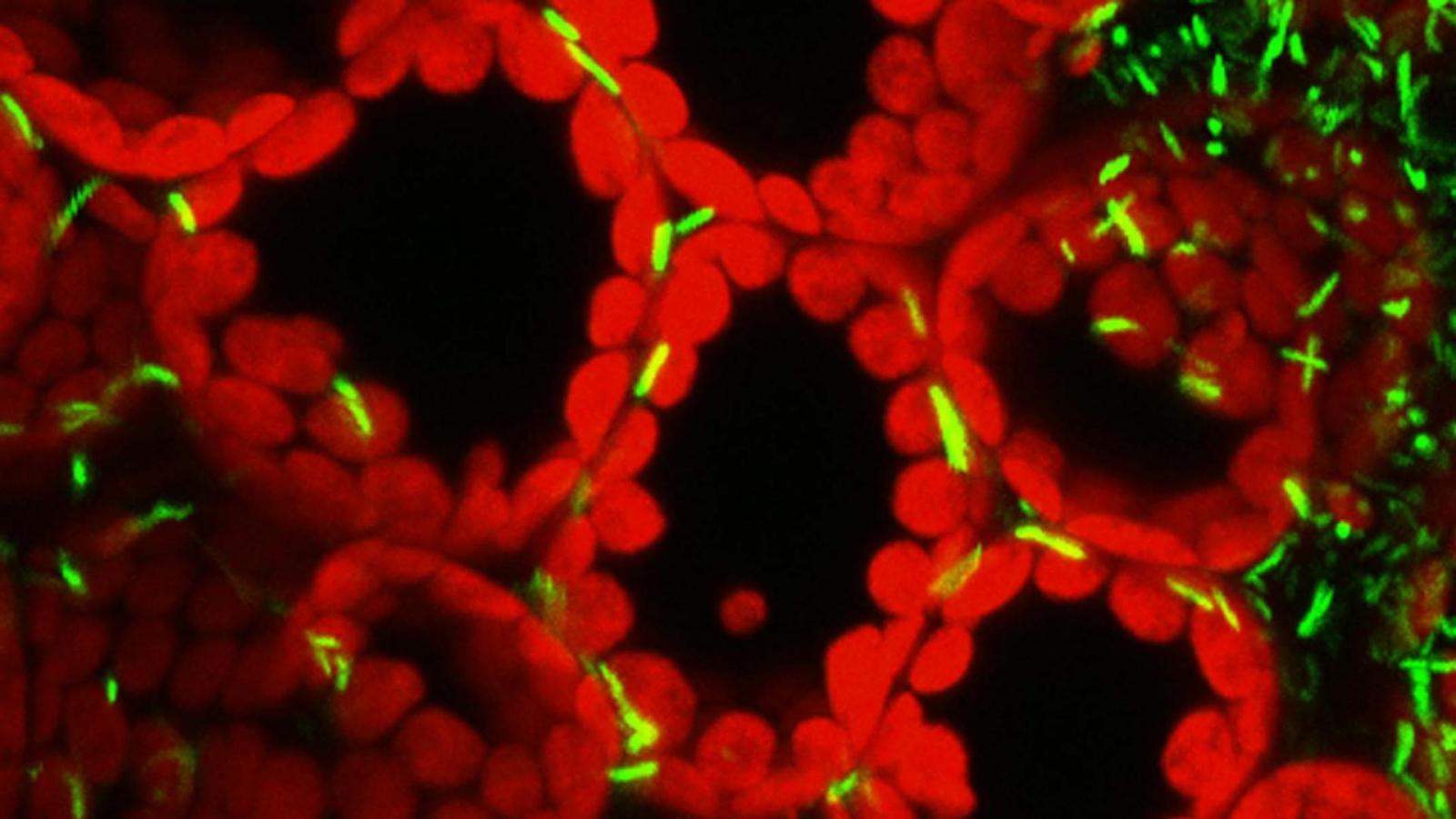
My research group focuses on understanding how the bacterial plant pathogen Pseudomonas syringae type III secretion system (T3SS) and the effector proteins it injects into plant cells disable plant immunity. Our early research on this system laid the groundwork for many subsequent discoveries by my laboratory. For example, my research group in collaboration with Alan Collmer's group found the type III apparatus genes were part of a pathogenicity island, which is a cluster of virulence genes that are horizontally acquired by the pathogen (Alfano et al., 2000). This, in turn, led to the discovery of several type III effector genes in the flanking regions of this pathogenicity island. My research group also developed several useful assays to determine whether a candidate type III effector was secreted in culture and/or injected into plant cells by the P. syringae T3SS (van Dijk et al., 1999). These discoveries later helped us identify other type III effectors from P. syringae strains as their genomes became available. Among our most significant accomplishment was the identification of common biochemical characteristics in the N-termini secretion signals of P. syringae type III effectors (Petnicki-Ocwieja et al., 2002). This made it relatively easy to identify type III effector genes in P. syringae strains by searching for genes downstream of conserved type III promoters that encoded products with N-termini characteristics resembling type III effectors.Since discovery of the complete inventory of P. syringae pv. tomato DC3000 (the strain we work with the most) type III effectors, our research has focused on determining their activities and targets inside plant cells. We were one of the first groups to demonstrate that individual plant pathogen type III effectors can suppress plant immunity (Jamir et al., 2004, Guo et al., 2009). Subsequently we performed genome-wide screens and found the majority of the DC3000 type III effectors were capable of suppressing immune responses (Jamir et al., 2004, Guo et al., 2009). A primary focus has been on identifying enzymatic activities and plant targets of type III effectors. For example, we found that the HopAO1 type III effector (formally known as HopPtoD2) is an active protein tyrosine phosphatase (Espinosa et al., 2003). We also found that the type III effector HopU1 is a mono-ADP-ribosyltransferase that can ADP-ribosylate several plant RNA-binding proteins (Fu et al., 2007). One of these RNA-binding proteins, GRP7, is involved in plant immunity because Arabidopsis plants lacking it are more susceptible to P. syringae and produce reduced immune responses compared to similarly challenged wild type plants. Because type III effectors are targeting plant immunity, we view them as important molecular tools for the identification new components of innate immunity that may not be identifiable using other approaches.
Current Research Goals
Identification of activities and targets of plant pathogen effectors
A major goal of my research group is to understand the roles effectors have inside plant cells. We believe that by continuing to focus on pathogen effectors, we will remain well positioned to identify new components of innate immunity. Further, there is a reasonable likelihood that these findings will represent translational research that will lead to novel biotechnological strategies to make crops more resistant to biotic stresses. Novel bioinformatic approaches to identify effector activities and plant targets are becoming increasingly important and, therefore, we are relying more and more on these approaches to identify their activities and targets inside plant cells. We are currently working on identifying plant targets and enzymatic activities of several DC3000 type III effectors.
Understanding how plant pathogen effectors cross the plant plasma membrane to gain entry into plant cells
In contrast to animal pathogens, plant pathogens need to breech the an additional barrier — the plant cell wall — in order to inject type III effectors into host cells. Thus, the injection or translocation of type III effectors into plant cells is likely quite different than in animal pathogens. We have focused on the P. syringae HrpK1 protein to begin to understand how bacterial T3SSs inject proteins into plant cells. HrpK1 is required for P. syringae to inject type III effectors into plant cells (Petnicki-Ocwieja et al., 2005). Also important in this process is the HrpZ1 harpin and other harpins. These proteins (HrpK1 and the harpins) likely make-up the “translocon” that forms pores in plant plasma membrane allowing type III effectors to gain entry into plant cells. Bacterial pathogens of animals have translocons made up of two translocator proteins (YopB and YopD in the prototypical Yersinia) and a tip protein (LcrV in Yersinia) which as it name suggests stays at the tip of the T3SS appendage (called the needle in most animal pathogens) where it binds to the translocators and probably positions them such that they are able to form a pore in the plasma membrane. HrpK1 has weak similarity to YopB, however, the plant pathogen translocon seems fundamentally different since there are no proteins similar to YopD or LcrV. We are interested in identifying the components of the translocon in P. syringae because it will help us get a better idea how the plant pathogens organize their translocons and how it translocates proteins across the plant cell wall and plasma membrane. Understanding how the translocon functions may lead to biotechnological strategies to inhibit translocation, which would prevent disease by bacterial pathogens dependent on T3SSs.
Understanding how the N-terminal secretion signals and/or type III chaperones are recognized by the T3SS apparatus
We are continuing to study how type III secretion signals and chaperones function to allow the T3SS apparatus recognize proteins that travel the type III secretion pathway. We hope to identify the order of secretion of type III-secreted proteins. The secretion hierarchy is probably critical to suppressing the plant innate immune system. Type III secretion signals, type III chaperones, and other control factors probably play major roles in determining which type III effectors are injected early and late in the translocation process. We have already identified one control factor in this process, the HrpJ protein. This is a type III secreted protein that needs to be present in the bacterial cell for HrpZ1 to be secreted and, therefore, acts as a control protein for secretion (Fu et al., 2006). In other T3SSs, type III chaperones bind to the type III ATPase, which is on the cytoplasmic side of the type III apparatus. Thus, this ATPase acts as a docking station for type III effector-chaperone pairs facilitating effector secretion. We are determining whether P. syringae type III chaperones play similar roles.
Other factors important for P. syringae pathogenesis of plants
We are initiating many other projects on P. syringae and plant immunity. One project that we are developing has to do with how the plant innate immune response is successful in blocking bacterial pathogen infections. When plant immunity is triggered, the plant produces many immune responses including reactive oxygen species, callose and cellulose, pathogen-related proteins, and a programmed cell death known as the hypersensitive response (HR). However, it is not known which output(s) is (are) effective at stopping bacterial infections. We are taking a genetic and cell biology approach to determine the extent that different plant mutants that lack specific immune responses are altered in their ability to contain bacterial pathogens.P. syringae has many other potential virulence factor genes that have been identified in its genome. We are determining the extent that they are important in pathogenesis by making bacterial mutants and testing how well they multiple in plants and produce disease symptoms. The virulence factors we are currently studying are those that relieve oxidative stress and those that produce different plant hormones. While the T3SS is critical to pathogenesis many of these other factors play key, perhaps critical, roles in the multfactorial process bacteria use to infect eukaryotes.